იონოტროპულ რეცეპტორთა სტრუქტურა
რეცეპტორები სპეცილიზებული ცილებია რომლებიც სასიგნალო სისტემების ინფორმაციას აღიქვამენ და შემდეგ უშუალოდ თუ სპეციალური ტრანსდუცერების საშუალებით გადასცემენ ბიოლოგიურ ინფორმაციას უჯრედს. განასხვავებენ ორი ტიპის იონოტროპულ და მეტაბოტროპულ რეცეპტორებს.იონოტროპული რეცეპტორები მეტაბოტროპულისაგან განსხვავებით სწრაფ უჯრედულ პასუხს იწვევს რადგან ისინი იონურ არხებს ქმნიან მათი აქტივაცია მემბრანათაშორის იონთა გადანაწილებას და შესაბამისად მოქმედების პოტენციალის ცვლილებას იწვევს რასაც სულ რაღაც მილიწამებიდან წამებამდე დრო სჭირდება ამიტომ იონოტროპულ რეცეპტორებს სწრაფი პასუხის რეცეპტორებს უწოდებენ არსებობს მეორე ტიპის რეცეპტორებიც რომლებიც ლიგანდის დაკავშირების შემდეგ კონფორმაციას იცვლის და G ცილის აქტივაციას განაპირობებს რომელიც თავის მხრივ ფერმენტ ადენილციკლაზას აქტივაციას განაპირობებს და მეორადი შუამავლის CAMP ( ციკლური ადენოზინმონოფოსფატის) დაგროვებას განაპირობებს.რაც უჯრედულ პასუხს განაპირობებს რაც დიდ დროს საჭიროებს ამიტომ ამ რეცეპტორებს ნელი მეტაბოტროპული რეცეპტორები ეწოდება. იონოტროპული რეცეპტორები როგორც წესი ჰომოლოგიური ცილებია ისინი ხუთი სუბერთეულისაგან შედგება ხოლო თითოეული სუბერთეული ოთხი დომენისაგან.
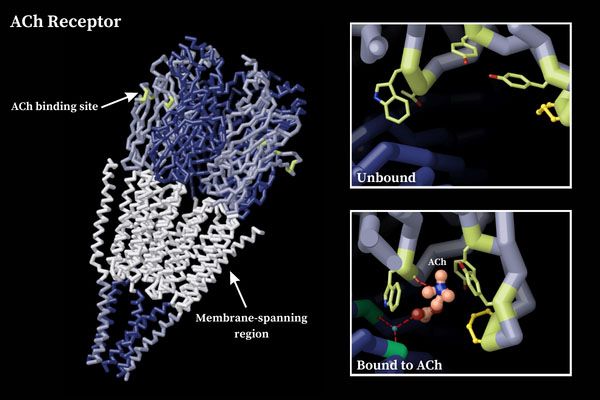
მათგან ყველაზე უკეთ შესწავლილი აცეტილქოლინის ნიკოტინური რეცეპტორია რომელიც ნატრიუმის იონურ არხს წარმოადგენს რეცეპტორის დიამეტრი 165 A-ია ხოლო ნაპრალის დიამეტრი 25-30 A. ლიგანდდამაკავშირებელი ცენტრი წარმოიქმნება გლიკოზილირებადი ნაშთებისა და ცისტეინის კავშირების საშუალებით,დომენების უჯრედგარეთა მხარეს N ბოლოზე. იონური არხის შიდა მხარე დატვირთულია უარყოფითად დამუხტული ამინომჟავური ნაშთებით რაც წარმოქმნის იონ-სელექტიურ ფილტრს და ატარებს მხოლოდ დადებითად დამუხტულ იონებს. თუმცა უნდა აღინიშნოს ისიც რომ არხის დიამეტრი საკმაოდ დიდია და წარმოადგენს წყლით სავსე ფორას რაც ამცირებს მის სელექტიურობას და საშუალებას იძლევა არხი გადაკვეთონ ისეთმა იონებმა როგორიცაა ტეტრამეთილამონიუმი ან სხვა მცირე ზომის დადებითი მუხტის მქონე იონები ან თუნდაც მცირე ზომის მოლეკულები. თუმცა არხში გავლა შესაძლებელია მხოლოდ და მხოლოდ დადებითი მუხტის მქონე იონებისათვის. აცეტილქოლინის დაკავშირების უნარი მხოლოდ და მხოლოდ მის ალფა სუბერთეულს გააჩნია და მისი დაკავშირების შემდეგ იონური არხი იხსნება რასაც მემბრანის დეპოლარიზაცია მოჰყვება. აღნიშნული რეცეპტორები ძირითადად გვხვდება ჩონჩხის კუნთებში ცნს-ში და თირკმელზედა ჯირკვლის ტვინოვან ნივთიერებაში.
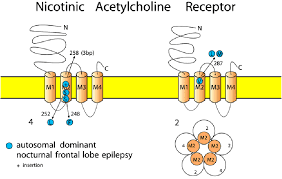
რაც შეეხება გლიცინის რეცეპტორს და თავად გლიცინს ეს ნეიროტრანსმიტერი ზურგის ტვინში გვხვდება და შემაკავებელ როლს ასრულებს. მისი რეცეპტორები ქლოორის იონს ატარებს და მემბრანის ჰიპერპოლარიზაციას იწვევს დეფიციტი შესაძლებელია ისეთ დაავადებებთან იყოს დაკავშირებული როგორიცაა ეპილეფსია.
რაც შეეხება გაემ-ის რეცეპტორს არსებობს ამ რეცეპტორის სამი ქვეტიპი გაემ-A გაემ-B გაემ-c
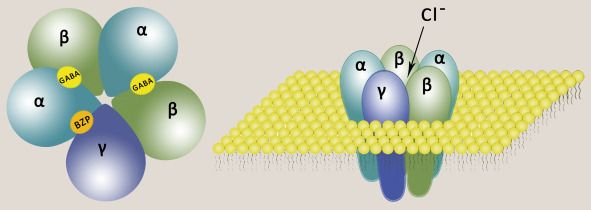
გაემ-A და გაემ-c აცეტილქოლინის ნიკოტინური რეცეპტორების მსგავსად ლიგანდდამაკავშირებელი ცენტრი რეცეპტორის N ბოლოზე მდებარეოობს და მის წარმოქმნაში ისევე როგორც აცეტილქოლინის რეცეპტორის შემთხვევაში მონაწილეობას გლიკოზილირებადი ნაშთები იღებს. რეცეპტორის შიდა მხარე დადებითად არის დამუხტული. გაემ A რეცეპტორს საინტერესოს რამოდენიმე ფაქტი ხდის. ამ რეცეპტორს გააჩნია ბენზოდიაზეპინების, ბარბიტურატების, სტეროიდული ჰორმონების დამაკავშირებელი უბანი და პიკროტოქსინის უბანი. ამ რეცეპტორს გააჩნია ფუროსემიდის უბანიც. ბენზოდიაზეპინები და ბარბიტურატები ფართოდ გამოყენებადი ანქსიოლიზური საშუალებებებია თუმცა ბენზოდიაზეპინებისაგან განსხვავებით ბარბიტურატები მალე იწვევს მიჩვევას და მათი თერაპიული ინდექსი ძალიან დაბალია რამაც შეიძლება ზედოზირება გამოიწვიოს. გაემ რეცეპტორის მრავალი სუბერთეულია კლონირებული ექვსი α, ოთხი β, სამი γ, ერთი φ და ა.შ ეს რეცეპტორები ცნს-ის სხვადასხვა რეგიონში შეიძლება განსხვავებული სუბერთეულოვანი
შემადგენლობით შეგვხვდეს შესაბამისად მათი მგრძნობელობა ერთი და იმავე მედიკამენტისადმი შეიძლება სრულიად განსხვავდებოდეს. რაც შეეხება გაემ-B რეცეპტორს ის მეტაბოტროპულია და ხსნის კალიუმის არხს რაც იწვევს მემბრანის ძლიერ ჰიპერპოლარიზაციას.
გლუტამატი მთავარი ამაგზნებელი რეცეპტორია ცნს-ში მას გააჩნია როგორც მეტაბოტროპული ისე იონოტროპული რეცეპტორები.იონოტროპული რეცეპტორები იონოტროპული რეცეპტორების დაყოფა შესაძლებელია სამ ჯგუფად ესენია ნმდა ამპა და კაინატის რეცეპტორები სხვა ზემოხსენებული რეცეპტორებისაგან განსხვავებით ნმდა რეცეპტორი ტეტრამერია და მას გააჩნია დისოციაციური ანესთეტიკების, მაგნიუმის დამაკავშირებელი, პოლიამინების სპერმინისა და სპერმიდინის დამაკავშირებელი უბანი თუთიისა და გლიცინის დამაკავშირებელი უბნები.ეს რეცეპტორი ატარებს დიდი რაოდენობით კალციუმის იონებს. კალციუმის დიდი კონცენტრაცია კი ციტო და ნეიროტოქსიურია,სწორედ ამიტომ იონური არხის გახსნას რამოდენიმე წინაპირობა ახლავს თან. საჭიროა მემბრანის ნაწილობრივი დეპოლარიზაცია ამისათვის აუცილებელია ამპა რეცეპტორების წინასწარი ძლიერი აქტივაცია. ნმდა რეცეპტორების აქტივაციისათვის აუცილებელია გლიცინის თანაარსებობაც თუმცა უჯრედში ყოველთვის არის ნმდა აქტივაციისათვის გლიცინის საკმარისი კონცენტრაცია.ნმდა რეცეპტორის აქტივაციის შემდეგ მას მოსცილდება მაგნიუმის იონები და რეცეპტორი იწყებს დიდი რაოდენობით კალციუმის იონების გატარებას მას სიგნალის ამპლიფიცირების ძლიერი უნარი აქვს. მისი ერთ-ერთი ფარმაკოლოგიურად ყველაზე საინტერესო უბანი დისოციაციური ანესთეტიკების უბანია მრავალი საანესთეზიო საშუალება მოქმედებს ამ უბანზე მათ შორის კეტამინიც. ეს უბანი რეცეპტორის შიდა მხარესაა ლოკალიზებული მისი ხანგრძლივი ბლოკადა შიზოფრენიის მსგავს ქცევას და ჰალუცინაციებს იწვევს თუმცა ეს უბანი კლინიკურად მაინც ძალზე საინტერესო რჩება რადგან მისი ხანმოკლე ბლოკადა აფერხებს კალციუმის ნეიროტოქსიურ ეფექტებს და ნეიროდეგენერაციულ პროცესებს შემუშავებულია მედიკამენტი მემანტინი რომელიც აღნიშნული მექანიზმით მოქმედებს და საკმაოდ ეფექტურია ალცჰეიმერის დაავადების მქონე პაციენტებში.აუცილებლად აღსანიშნავია ისიც,რომ ნმდა რეცეპტოორის აქტივაცია წამყვან როლს ასრულებს დასწავლის პროცესებში.
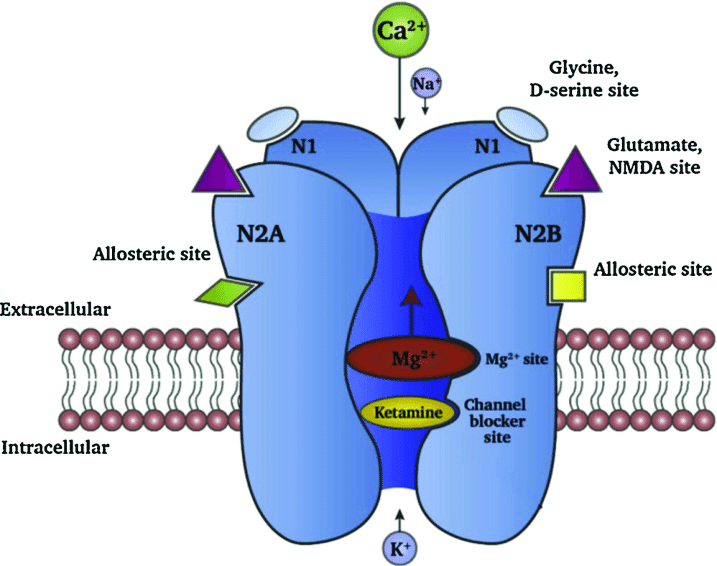
რაც შეეხება ამპა რეცეპტორს ის ოთხი დომენისაგან შედგება და თითოეული დომენი ოთხი სუბერთეულისაგან რომლებიც იონური არხის ფორმირებაში იღებენ მონაწილეობას. ამ რეცეპტორს გააჩნია 130000 კდ-ის მქონე რეგულატორული სუბერთეულიც. რაც შეეხება კაინატის რეცეპტორებს ესენი ორი სხვადასხვა ოჯახის რეცეპტორებისაგან შედგება GluR5-7 და KA-1 და KA-2. სუბერთეულები წარმოქმნის კაინატის დამაკავშირებელ უბნებს ხოლო GluR5-7 რეცეპტორის ფორმირებაში მონაწილეობს. GluR5-7 სუბერთეულების გარეშე ისინი უჯრედის მემბრანაზე ვერ ექსპრესირდება და ენდოპლაზმურ რეტიკულუმში რჩება.
არსებობს იონოტროპულ რეცეპტორთა კიდევ ერთი ჯგუფი პოტენციალმგრძნობიარე არხები რომლებიც ოთხი დომენისაგან შედგება მათი მეოთხე დომენი დანარჩენებისაგან იმით განსხვავდება რომ ის მდიდარია დადებითად დამუხტული ამინომჟავებით ლიზინითა და არგინინით ისინი ჰიდროფობურ ამინომჟავებთან ურთიერთქმედებით წარმოქმნიან დადებითი ამინომჟავების მთელ სვეტს პოტენციალის ცვლილების შემდეგ ეს დომენი კონფორმაციას იცვლის და აქტივდება. პოტენციალმგრძნობიარეა კალციუმის არხებიც რომელიც ანტიჰიპერტენზიული პრეპარატების სამიზნეა ისეთი პრეპარატები როგორებიცაა ამლოდიპინი დილტიაზემი და ვერაპამილი სწორედ ამ იონურ არხთან ურთიერთქმედებენ. დიჰიდროპირიდინების უბანი იონური არხის გარეთა მხარესაა მოთავსებული ხოლო ფენილალკილამინებისა დაბენზოტიაზეპინების უბნები ამ არხების შიგნით მდებარეობს. აღსანიშნავია რომ არსებობს კალციუმდამოკიდებული კალიუმის არხებიც რომლებიც მონაწილეობენ უამრავი ჰორმონის მათ შორის ინსულინის სეკრეციაშიც.
გამოყენებული ლიტერატურა :
ნ.ნაცვლიშვილი დ. მიქელაძე " უჯრედული პროცესების რეგულაციის ძირითადი პრინციპები "